
ในโลกที่ตระหนักถึงความยั่งยืนด้านสิ่งแวดล้อม และต้องการประสิทธิภาพการใช้พลังงานมากขึ้น พลังงานแสงอาทิตย์จึงกลายเป็นทางออกที่หลาย ๆ คนต้องการ แต่ต้นทุนการติดตั้งแผงโซลาร์เซลล์ยังคงเป็นอุปสรรคสำคัญเพราะมีราคาที่สูง

โรงไฟฟ้าพลังงานน้ำ ประเภทโรงงานที่ผลิตไฟฟ้าให้ใช้งาน โรงไฟฟ้าพลังงานน้ำ ประเภทโรงงานที่ผลิตไฟฟ้าให้ใช้งาน
น้ำเป็นทรัพยากรธรรมชาติที่สำคัญต่อการเลี้ยงชีวิต ใช้เพื่อบริโภคและอุปโภค รวมถึงการผลิตไฟฟ้า คนไทยจึงมีแสงสว่างจากไฟฟ้าให้ใช้งานมาเป็นเวลาเกินกว่าครึ่งศตวรรษ การผลิตนั้นจะมาจากโรงไฟฟ้าพลังงานน้ำซึ่งปัจจุบันมีอยู่หลายประเภท โดยสามารถกล่าวรายละเอียดได้ในบทความนี้ให้ทุกคนรู้จักมากขึ้นกว่าเดิม โรงไฟฟ้าพลังงานน้ำ ให้อะไรบ้างแก่ชาวไทย พลังงานน้ำถูกนำมาใช้ประโยชน์ตั้งแต่อดีตถึงปัจจุบัน ขึ้นอยู่กับแต่ละคนว่านำน้ำมาทำอะไรบ้าง แต่หากให้พูดถึงการผลิตไฟฟ้านั้น ต้องมีเครื่องกำเนิดไฟฟ้าเป็นอุปกรณ์หลักซึ่งตัวเครื่องนี้ถูกพัฒนาเมื่อปลายศตวรรษที่ 19 ส่วนประเทศไทยก็จริงจังเรื่องการผลิตไฟฟ้าในปี 2507 ดำเนินการ ณ เขื่อนภูมิพล จังหวัดตาก เวลาผ่านไปก็ได้มีโรงไฟฟ้าพลังงานน้ำเกิดขึ้นมากกว่าเดิมซึ่งจัดอยู่ในประเภทต่าง ๆ ไม่ว่าจะโรงงานไหนล้วนผลิตไฟฟ้าจากพลังงานน้ำจึงไม่ส่งผลกระทบต่อสิ่งแวดล้อมและลดการปล่อยก๊าซคาร์บอนไดออกไซด์อันเป็นสาเหตุให้เกิดภาวะโลกร้อน การผลิตยืดหยุ่นสูงอีกทั้งต้นทุนต่ำ ไฟฟ้าที่ได้มีปริมาณมาก สามารถใช้งานได้อย่างทั่วถึง ประเภทโรงไฟฟ้าพลังงานน้ำ มีอะไรบ้าง ในส่วนของประเภทโรงไฟฟ้าพลังงานน้ำ ประเทศไทยมีอยู่ด้วยกันทั้งหมด 3

ข้อควรรู้ของขับขี่โดยไม่มีประกันรถยนต์ในประเทศไทยข้อควรรู้ของขับขี่โดยไม่มีประกันรถยนต์ในประเทศไทย
การขับขี่โดยไม่มีประกันรถยนต์ในประเทศไทยเป็นการกระทำที่ผิดกฎหมายและสามารถส่งผลกระทบทางกฎหมายและทางการเงินต่อผู้ขับขี่ได้ ในบทความนี้ เราจะมาสำรวจถึงผลกระทบที่อาจเกิดขึ้นจากการขับขี่โดยไม่มีประกันรถยนต์กัน ค่าปรับและความรับผิดชอบที่อาจเกิดขึ้น การขับขี่โดยไม่มีประกันรถยนต์สามารถนำไปสู่การถูกปรับและความรับผิดชอบที่อาจเกิดขึ้นสำหรับผู้ขับขี่ หากเกิดอุบัติเหตุ ผู้ขับขี่อาจต้องรับผิดชอบค่าใช้จ่ายทั้งหมดที่เกี่ยวข้อง ไม่ว่าจะเป็นค่ารักษาพยาบาล ค่าซ่อมแซมรถ หรือค่าชดเชยสำหรับผู้ที่ได้รับบาดเจ็บในเหตุการณ์ และถ้าผู้ขับขี่ไม่มีประกันรถยนต์ 3+ ราคาที่เหมาะสม ความรับผิดชอบทางการเงินนี้อาจส่งผลกระทบอย่างรุนแรงต่อสภาพการเงินของผู้ขับขี่ การเปลี่ยนแปลงการใช้ประกันรถยนต์ในสังคมไทย ก่อนหน้านี้ ผู้ขับขี่อาจเลือกที่จะขับขี่โดยไม่มีประกันรถยนต์ 3+ ราคาที่เข้าถึงได้ง่าย แต่หลังจากที่ได้มีความเข้าใจเกี่ยวกับผลกระทบทางกฎหมายและการเงินที่อาจเกิดขึ้น ผู้ขับขี่หลายคนเป็นส่วนใหญ่ในสังคมได้ตัดสินใจที่จะซื้อประกันรถยนต์เพื่อปกป้องตัวเองจากความเสี่ยงที่อาจเกิดขึ้นมากขึ้น การป้องกันความเสี่ยงด้วยประกันรถยนต์ การมีประกันรถยนต์ 3+ ราคาที่เหมาะสมเป็นวิธีที่ดีที่สุดในการป้องกันความเสี่ยงที่อาจเกิดขึ้นจากการขับขี่ ไม่ว่าจะเป็นค่าใช้จ่ายในการซ่อมแซมรถ ค่ารักษาพยาบาล หรือค่าชดเชยสำหรับผู้ที่ได้รับบาดเจ็บ การมีประกันรถยนต์ที่เหมาะสมจะช่วยปกป้องผู้ขับขี่จากความเสี่ยงทางการเงินและทางกฎหมายที่อาจเกิดขึ้นได้ การปฏิบัติตามกฎหมายและการปกป้องตัวเอง

หัวพอต Time Pod แบรนด์น้องใหม่ โดดเด่น มาแรงน่าสนใจตั้งแต่แพคเกจหัวพอต Time Pod แบรนด์น้องใหม่ โดดเด่น มาแรงน่าสนใจตั้งแต่แพคเกจ
Time Pod หัวพอตไฟฟ้าแบรนด์น้องใหม่ ที่พึ่งเข้ามาตีตลาดในบ้านเราได้ไม่นาน ปรากฏมีเสียงตอบรับที่ดีเกินคาด จะบอกว่าน่าสนใจตั้งแต่แพตเกจ ดีไซน์แพคเกจมาโดดเด่นเร้าใจ แอบแฝงไปด้วยความคลาสสิค ผสมผสานเข้ากันได้อย่างลงตัว หัวตอนสามารถใช้ร่วมกับพอตไฟฟ้าได้หลายค่าย ไม่ว่าจะเป็น Relx Infinity, Hues, INFY ปัจจุบันมีน้ำยาให้เลือกมากกว่า 10 กลิ่น Time Pod หัวพอตที่กำลังมาแรง ในปี 2023 ต้องยอมรีบเลยว่าTime Pod หัวพอตแบรนด์นี้มาแรงแซงโค้งแม้จะเป็นแบรนด์น้องใหม่ที่พึ่งเปิดตัวได้ไม่นานแต่ทำยอดขายได้ดีเกินคาด กลายเป็นหัวพอตที่ถูกพูดถึงในวงกว้าง หัวพอตเป็นแบบใสทำให้มองเห็นน้ำในข้างในได้ สังเกตได้ว่าน้ำยาจะหมดเมื่อไหร่ นอกจากนั้นยังไม่ต้องกังวลเรื่องน้ำยารั่วซึมด้วยทางค้ายออกแบบมาอย่างดี

เทคนิคเลือกซื้อบีบีกันอย่างไรไม่ให้โดนหลอกเทคนิคเลือกซื้อบีบีกันอย่างไรไม่ให้โดนหลอก
บีบีกัน หรือที่เรียกในอีกชื่อหนึ่งว่า AIRSOFT GUN หรือ ปืนอัดลมเบา เป็นเกมกีฬายิงปืนในรูปแบบใหม่ ที่มีความปลอดภัยต่อผู้เล่น และเป็นเกมกีฬาที่ได้รับความนิยมมากในหลายประเทศทั่วโลก อาทิ ญี่ปุ่น ไต้หวัน ฮ่องกง อังกฤษ อเมริกา ฯลฯ เป้าหมายของการใช้บีบีกันคือใช้งานกิจกรรมสันทนาการต่าง ๆ ใช้สำหรับแข่งขันเป็นกีฬาหรือเอาไว้ใช้สะสมสำหรับผู้ที่ชื่นชอบอาวุธปืนเนื่องจากการถือครองอาวุธปืนจริงอาจก่อให้เกิดอันตรายถึงชีวิตได้ ขณะที่ปืน BB Gun ไม่เป็นอันตรายต่อบุคคลรอบข้างแต่ถึงอย่างนั้นมันก็สร้างความเสียหายได้เล็กน้อยถึงปานกลาง วันนี้จะมาบอกเทคนิคเลือกซื้อบีบีกันอย่างไรไม่ให้โดนหลอกกันเลย แยกประเภทปืนให้ถูก บีบีกัน มีทั้งผลิตจากญี่ปุ่น ฮ่องกง ใต้หวัน และจีน

เคล็ดลับการเลือกประกันภัยรถยนต์ให้ได้ความคุ้มครองที่ถูกใจเคล็ดลับการเลือกประกันภัยรถยนต์ให้ได้ความคุ้มครองที่ถูกใจ
หากจะกล่าวถึงความสำคัญของประกันภัยรถยนต์แล้ว เชื่อเหลือเกินว่า เหล่าผู้ขับขี่ยานยนต์จำนวนไม่น้อยเลยที่มีองค์ความรู้ และเข้าใจเรื่องความสำคัญของมันเป็นอย่างดี เมื่อถึงเวลาที่จะต้องต่อประกันแต่ละครั้งจะต้องทำการเปรียบเทียบ ประกันรถยนต์ เพื่อให้ได้รับแผนความคุ้มครองที่ดีเสมอ แต่สำหรับผู้ขับขี่ที่ยังเป็นมือใหม่ อาจจะยังไม่เข้าใจมากนักว่าเพราะเหตุใด การทำประกันรถยนต์จึงสำคัญกับการขับขี่มากขนาดนั้น แล้วจำเป็นต้องเปรียบเทียบ ประกันรถยนต์ เพื่อดูความคุ้มครองด้วยหรือ? บทความนี้จะพามาหาคำตอบกัน ประกันภัยรถยนต์ช่วยคุณได้อย่างไร มันสามารถปกป้องคุณทางการเงินได้หากคุณประสบอุบัติเหตุ หากคุณทำให้เกิดอุบัติเหตุ ประกันรถยนต์สามารถช่วยจ่ายค่าเสียหายของผู้ขับขี่คนอื่นได้ และหากคุณคือผู้ที่ได้รับความเสียหายจากอุบัติเหตุ ประกันภัยรถยนต์สามารถช่วยจ่ายค่าซ่อมให้คุณได้ หากคุณกังวลว่าจะประสบอุบัติเหตุ การทำประกันรถยนต์จะช่วยให้คุณสบายใจได้ และหากคุณประสบอุบัติเหตุ ก็สามารถช่วยดูแลให้สถานการณ์กลับมาเป็นปกติให้ได้เร็วมากขึ้น ดังนั้น หากคุณกำลังคิดจะทำประกันรถยนต์ ก็สามารถพิจารณาความคุ้มครองต่าง ๆ เพื่อหาแผนประกันที่เหมาะสมกับการขับขี่รถยนต์ของคุณ โดยเลือกใช้บริการเปรียบเทียบ

การเปิดร้านเกี่ยวกับผลิตภัณฑ์สิ่งแวดล้อมดีอย่างไรการเปิดร้านเกี่ยวกับผลิตภัณฑ์สิ่งแวดล้อมดีอย่างไร
ในปัจจุบันนี้เรื่องของสิ่งแวดล้อมนั้นก็เป็นเรื่องหนึ่งที่ดีและสำคัญกับเราเองอย่างยิ่งเลย เราเองจึงควรที่จะต้องอย่ามองข้ามแล้วก็ให้ความสนใจกับเรื่องของสิ่งแวดล้อมด้วยจึงจะดีเพราะว่าในเรื่องของสิ่งแวดล้อมนั้นก็เป็นสิ่งที่จะช่วยทำให้โลกของเรานั้นเย็นขึ้นด้วยและที่สำคัญในเรื่องของสิ่งแวดล้อมนั้นจะช่วยทำให้เรายิ่งมีความสุขอีกด้วย เพราะว่าการที่เราเป็นคนรักสิ่งแวดล้อมเราก็อยากที่จะต้องหาผลิตภัณฑ์เกี่ยวกับสิ่งแวดล้อมมาขายด้วยเพราะว่าเราก็จะได้สนับสนุนให้คนอื่นๆนั้นให้ความสนใจกับเรื่องของสิ่งแวดล้อมด้วยและที่สำคัญในเรื่องของสิ่งแวดล้อมนั้นก็เหมือนกับเป็นจุดศูนย์รวมที่ให้คนที่สนใจในผลิตภัณฑ์ที่ไม่ทำร้ายธรรมชาตินั้นเข้ามาเลือกซื้อและเลือกใช้กันด้วย การที่เราเปิดร้านเกี่ยวกับธรรมชาตินี้นั้นก็จะมีในส่วนของซิลิโคนที่ไว้ใช้สำหรับใส่ของแทนอาหาร สิ่งนี้จะยิ่งเป็นสิ่งที่ดีอย่างมากที่จะช่วยทำให้เราได้ลดการใช้ถุงพลาสติก ลดในเรื่องของโฟม ลดในสิ่งที่จะทำให้ย่อยสลายช้าและทำให้ร้อนด้วย เรื่องนี้จึงเป็นเรื่องที่ดีอย่างมากเลย ดังนั้นในตอนนี้จึงมีคนหันกลับมาใช้ของที่ย่อยสลายกันง่ายเช่นจานกระดาษ หรือวัสดุที่ทำมาจากวัสดุธรรมชาตินั้นเองด้วย สิ่งเหล่านี้จึงจะเป็นเรื่องที่เราเองควรที่จะต้องให้ความสนใจกันอย่างที่สุดเพื่อสิ่งแวดล้อมของเราก็จะได้ย่อยสลายง่ายและไม่เป็นพิษอีกด้วย เรื่องนี้จึงเป็นเรื่องที่เราเองก็ควรที่จะต้องให้ความสนใจและอย่าละเลยไปเลยจึงจะยิ่งดีและที่สำคัญในเรื่องของสิ่งแวดล้อมนั้นเราก็จะต้องอย่าละเลยไปเลยจะได้ไม่ส่งผลเสียอีกด้วย ในเรื่องของธรรมชาตินั้นต้องบอกเลยว่าเป็นสิ่งที่ดีกับเราเองอย่างยิ่งเลยเพราะถ้าหากเรารู้จักที่จะใส่ใจแล้วก็จะช่วยทำให้เราได้มีโลกที่เย็นขึ้นเพราะว่าโลกของเราในตอนนี้นั้นมีอากาศที่ร้อนอย่างมากเลยดังนั้นเราจึงควรที่จะต้องอย่าละเลยหรือมองข้ามไปเลยจึงจะดีที่สุดเพราะหากเราได้ให้ความสำคัญแล้วนั้นก็จะทำให้เรายิ่งมีความสุขยิ่งขึ้นด้วย ในเรื่องของโลกร้อนนั้นเราอาจจะพบเจอกันอย่างมากมายในทุกๆวันและเรื่องนี้จึงเป็นเรื่องที่เราเองก็ควรที่จะต้องให้ความสำคัญมากๆด้วยจึงจะยิ่งดีอย่างน้อยในเรื่องของโลกร้อนนั้นเราก็มีส่วนที่จะสามารถช่วยลดในเรื่องของความร้อนได้ด้วย สิ่งนี้จึงเป็นเรื่องที่เราเองก็จะต้องอย่าละเลยไปเลยจึงจะดีที่สุด

ผลผลิตที่เกี่ยวกับสิ่งแวดล้อมมีอะไรน่าสนใจบ้างผลผลิตที่เกี่ยวกับสิ่งแวดล้อมมีอะไรน่าสนใจบ้าง
ในปัจจุบันนี้เรื่องของสิ่งแวดล้อมนั้นก็เป็นเรื่องหนึ่งที่ดีและสำคัญกับเราเองอย่างมากเลยเพราะว่าในเรื่องของผลผลิตที่เกี่ยวกับสิ่งแวดล้อมนั้นจะช่วยทำให้เรายิ่งมีความเป็นอยู่หรือมีสิ่งแวดล้อมที่ดียิ่งขึ้นด้วยและที่สำคัญในเรื่องของสิ่งแวดล้อมนั้นจะช่วยทำให้เรานั้นยิ่งมีความสุขอย่างยิ่งด้วย และในตอนนี้เรื่องของการอนุรักษ์ในสิ่งแวดล้อมนั้นก็เป็นเรื่องที่ดีเนื่องจากว่าหากเราได้ให้ความสนใจแล้วนั้นก็จะช่วยทำให้เรายิ่งมีความสุขกันอย่างที่สุดอย่างน้อยการที่เราผลิตสิ่งแวดล้อมนั้นก็จะช่วยที่จะทำให้เกิดการลดโลกร้อนด้วยในตอนนี้ก็มีผลิตภัณฑ์ที่เกี่ยวกับสิ่งแวดล้อมอย่างมากมายเลย ในเรื่องของผลผลิตเกี่ยวกับสิ่งแวดล้อมนั้นก็จะมีในส่วนของผลิตภัณฑ์แชมพูที่มาจากสมุนไพรหรือกากมะพร้าวที่ทำออกมาเป็นใยขัดฟองน้ำด้วยเพราะในเรื่องของสิ่งแวดล้อมนั้นก็เป็นเรื่องที่ดีที่จะช่วยทำให้เรานั้นได้ใช้ผลิตภัณฑ์ที่มาจากสิ่งแวดล้อมด้วย และทุกๆอย่างในตอนนี้เรื่องของสิ่งแวดล้อมนั้นก็เป็นเรื่องที่เราเองก็จะต้องอย่ามองข้ามผ่านเลยจึงจะดีที่สุดเพราะถ้าหากเรามองข้ามผ่านไปแล้วอาจจะไม่ใช่เรื่องที่ดีอีกด้วยและแน่นอนว่าในเรื่องของสิ่งแวดล้อมนั้นจะช่วยทำให้เรานั้นได้สัมผัสกับธรรมชาติได้อย่างมากมายที่สุดด้วย เราจึงควรที่จะต้องให้ความสนใจและอย่าละเลยไปเลยจึงจะยิ่งดีและแน่นอนว่าในเรื่องของสิ่งแวดล้อมนอกจากจะเป็นของใช้ส่วนตัวแล้วก็จะยังเป็นในส่วนของผลิตภัณฑ์ในบ้านด้วยซึ่งทั้งหมดนี้ก็เป็นเรื่องที่เราเองก็จะต้องอย่ามองข้ามเลยอย่างน้อยสิ่งที่เราได้สัมผัสธรรมชาตินั้นก็จะส่งผลที่ดีอีกด้วย และแน่นอนว่าในเรื่องของการอนุรักษ์ในเรื่องของสิ่งแวดล้อมนั้นก็มักจะเป็นเรื่องที่ดีอย่างที่สุดและที่สำคัญในเรื่องของสิ่งแวดล้อมนั้นยิ่งเราได้ใช้ของที่ดีได้มากเท่าไหร่ก็จะช่วยทำให้เราได้ยิ่งมีความผ่อนคลายอย่างที่สุดเลยและที่สำคัญในเรื่องของสิ่งแวดล้อมจะช่วยทำให้เรายิ่งเป็นคนที่ดียิ่งขึ้นด้วยเพราะเราก็จะได้มีความสุขที่ได้เข้าใจในธรรมชาติอย่าแท้จริงอีกด้วย เรื่องของสิ่งแวดล้อมในตอนนี้จึงเป็นเรื่องที่เราเองก็ควรที่จะต้องอย่าละเลยหรือมองข้ามไปเลยจึงจะดีที่สุดเพราะถ้าหากเราละเลยหรือมองข้ามไปแล้วอาจจะส่งผลกระทบกับตัวเราเองด้วยและแน่นอนว่าในเรื่องนี้จึงเป็นเรื่องที่เราเองก็จะต้องใส่ใจให้มากๆด้วยเพราะในเรื่องของผลิตภัณฑ์ที่เกี่ยวกับสิ่งแวดล้อมนั้นจะช่วยทำให้เรายิ่งได้ใช้อย่างไม่มีสารเคมีเข้ามาสะสมในร่างกายได้อีกเช่นกัน
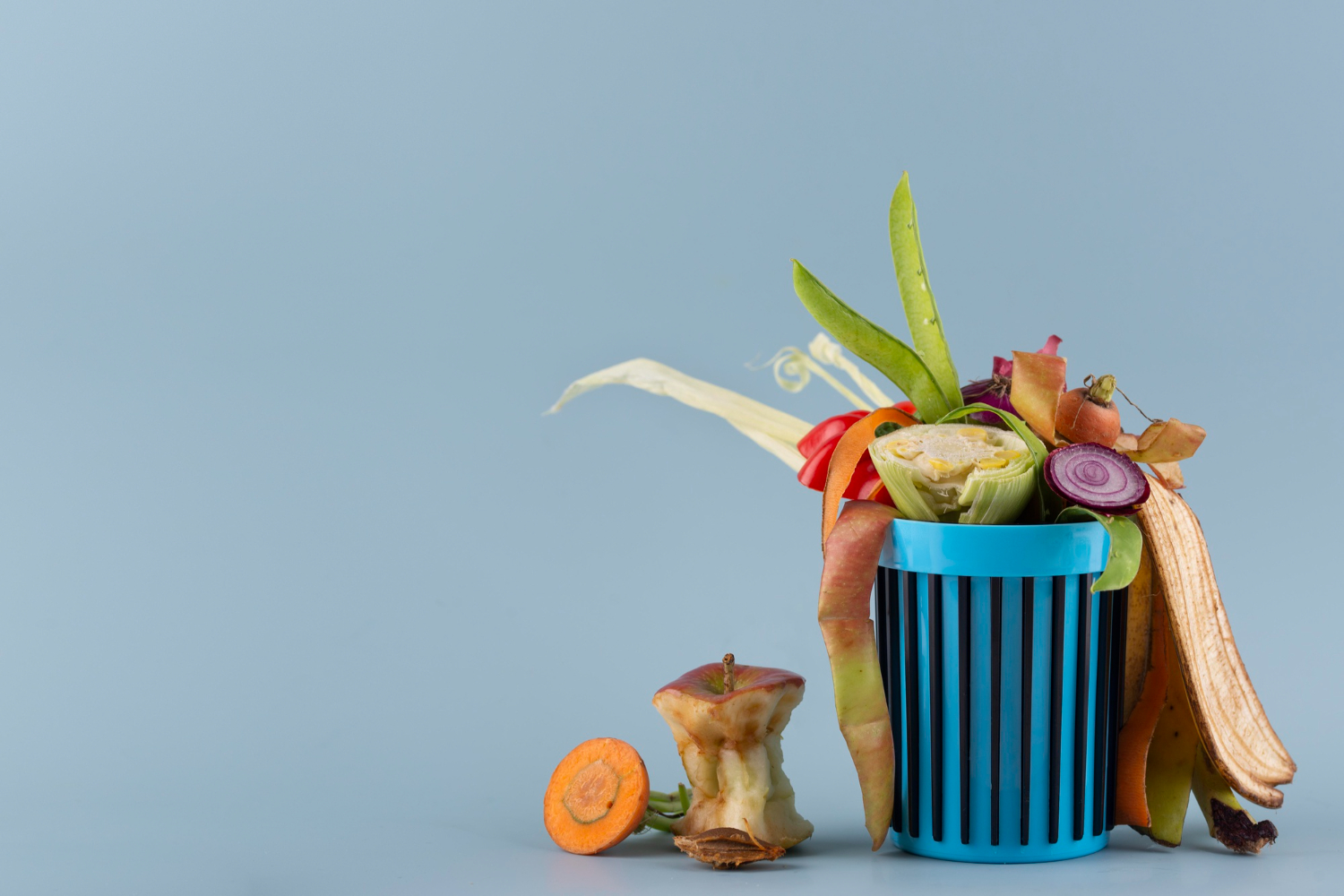
การช่วยกันแยกเศษอาหารเป็นเรื่องดีอย่างไรการช่วยกันแยกเศษอาหารเป็นเรื่องดีอย่างไร
ในปัจจุบันนี้ต้องบอกเลยว่าเรื่องของสิ่งแวดล้อมนั้นเป็นเรื่องหนึ่งที่ดีและสำคัญกับเราเองอย่างที่สุดดังนั้นเรื่องของสิ่งแวดล้อมจึงเป็นเรื่องที่เราเองก็ควรที่จะต้องให้ความสนใจและอย่ามองข้ามไปเลยจึงจะดีที่สุดและที่สำคัญในเรื่องของสิ่งแวดล้อมนั้นอย่างในเรื่องของการแยกเศษอาหารนั้นก็จะเป็นเรื่องที่ดีที่จะช่วยทำให้เรานั้นได้ลดปริมาณของขยะได้อีกเช่นกัน เพราะเรื่องของเศษอาหารนั้นจะช่วยทำให้เราได้เป็นคนที่มีความสุขอย่างที่สุดเลยและที่สำคัญในเรื่องของสิ่งแวดล้อมนั้นเป็นเรื่องที่เราก็สามารถที่จะร่วมทำได้ดังนั้นเรื่องของการช่วยแยกเศษอาหารนั้นก็เป็นเรื่องที่ดีที่เราเองก็จะละเลยไปไม่ได้และหากเราละเลยไปแล้วนั้นอาจจะทำให้เราไม่มีความสุขอีกด้วย ทุกๆอย่างเป็นสิ่งที่ดีอย่างยิ่งดังนั้นเราจึงควรที่จะต้องอย่ามองข้ามผ่านไปเลยและที่สำคัญหากเราได้แยกเศษอาหารแล้วก็จะง่ายต่อการลดปริมาณของขยะด้วย เรื่องทั้งหมดนี้จึงเป็นเรื่องที่เราเองก็ควรที่จะต้องอย่าละเลยไปเลยจึงจะส่งผลที่ดีและแน่นอนว่าในเรื่องของการที่เราช่วยแยกเศษอาหารนั้นก็จะมีส่วนที่สำคัญที่จะช่วยทำให้เรามีความสุขอย่างที่สุด ในการที่เราจะแยกเศษอาหารนั้น อย่างน้อยเราก็จะถูกแยกภาชนะแล้วเรื่องนี้จึงเป็นเรื่องที่เราเองก็ควรที่จะต้องอย่าละเลยหรือมองข้ามไปเลยจึงจะยิ่งดีเพราะถ้าหากเรามองข้ามหรือละเลยไปแล้วนั้นอาจจะส่งผลกระทบให้กับตัวเราเองได้อย่างยิ่งด้วยและในตอนนี้เรื่องของการแยกเศษอาหารนั้นก็จะทำให้เราเป็นคนที่มีความสุขอย่างที่สุดและที่สำคัญหากเราได้ให้ความสนใจแล้วจะทำให้เรายิ่งลดปริมาณขยะได้อีกเหมือนกัน อะไรที่จะยิ่งช่วยทำให้เรานั้นมีความสุขและเกิดความสุขได้ก็จะต้องยิ่งทำเพราะว่าความสุขของเราก็คือความภูมิใจให้กับตัวเราเองได้อย่างมากมายเลย และแน่นอนว่าในเรื่องของการแยกเศษอาหารนั้นจะช่วยทำให้นั้นยิ่งดูดีมากขึ้นได้และเรื่องนี้เราก็ควรที่จะต้องช่วยลดปริมาณขยะได้เพียงแค่เราจะต้องหมั่นมีวินัยในการแยกเศษอาหารเพียงเท่านี้ก็จะช่วยทำให้เรามีความสุขอย่างที่สุดเลย เราจึงควรที่จะต้องอย่าละเลยหรือมองข้ามไปเลยจึงจะยิ่งดีเพราะในเรื่องของสิ่งแวดล้อมนั้นก็จะเป็นเรื่องที่จะช่วยทำให้เรานั้นยิ่งมีความสุขกันอย่างมากมายด้วยนั้นเอง